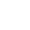中文





免疫组库测序在各种疾病研究中显示出良好的前景,如自身免疫性疾病、传染病、肿瘤的诊断、监测和治疗等,免疫组库测序。作为当下备受瞩目的前沿技术之一,正为我们揭开免疫系统奥秘的面纱。免疫组库测序不仅让我们能够深入了解身体对抗病原体的方式,更为理解免疫相关疾病和药物研发提供了新的视角。本文旨在向大家介绍什么是免疫组库测序,无论您是否对生物科学有着浓厚兴趣,本文都将为您打开一扇通往免疫学世界的门,让我们一起探寻人类抵御疾病的绝妙之处吧!
PART 01
免疫组库测序测的是什么?
免疫系统保护人体免受病原体的伤害,这种保护主要为大量淋巴细胞受体、T细胞受体(T cell receptor, TCR)和B细胞受体(B-cell receptor, BCR)诱导的免疫反应。免疫受体是抗原结合的重要结构域,共有七条链参与形成受体,T细胞包括由α链和β链编码的αβT细胞或由γ链和δ链编码的γδT细胞,B细胞受体由重链和κ链或重链和λ链编码,还有双α和双γT细胞,这些受体有助于确定抗原特异性。在任何特定时刻,一个人的免疫系统都由独特的TCR和BCR组成,这反映了免疫系统对毒素或外来物质的反应能力(图1) 。
免疫组库测序是指检测BCR及TCR,B细胞上负责识别并特异结合抗原的膜表面免疫球蛋白,我们称为BCR,BCR是由两条重链和两条轻链组成;T细胞上特异识别并特异结合抗原的分子为TCR,TCR分子也属于免疫球蛋白超家族, 由α、β或γ、δ两条肽链组成。编码BCR/TCR的4个基因簇V(可变区)、D(高变区)、J(编码蛋白连接v、c区的结合区)、C(恒定区) , B/T细胞通过V(D)J重组,使BCR/TCR基因序列极为丰富。V(D)J序列多样性的集合我们称为为免疫组库(Immune Repertoire, IR),通过高通量测序,可一次性获得大量免疫组库数据,从而实现免疫组库高通量测序。
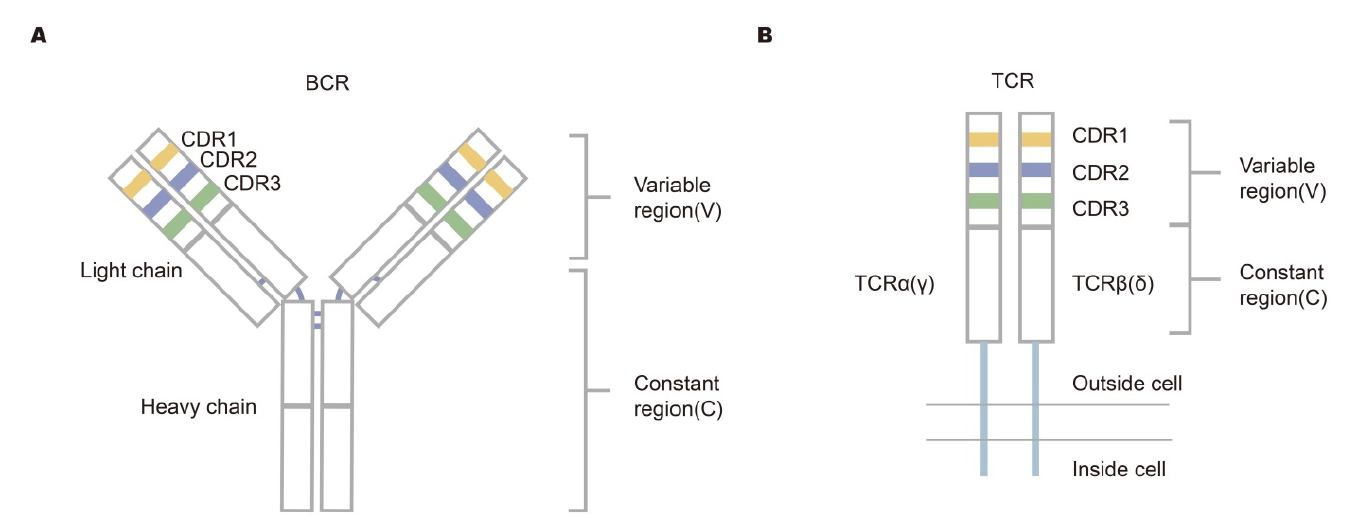
图1 抗原特异性淋巴细胞受体的结构和多样性[1]
注:上图显示了抗体的重链和轻链,它们通过二硫键连接(粗蓝线);跨细胞膜的TCR是由αβ链或γδ链组成的异二聚体。上图中由V(D)J基因组成的可变(V)区,V区由CDR区和框架区(FR)组成。CDR1、CDR2和CDR3显示为不同的颜色(分别为黄色、蓝色和绿色)。下部(白色区域)是一个保守的恒定区域
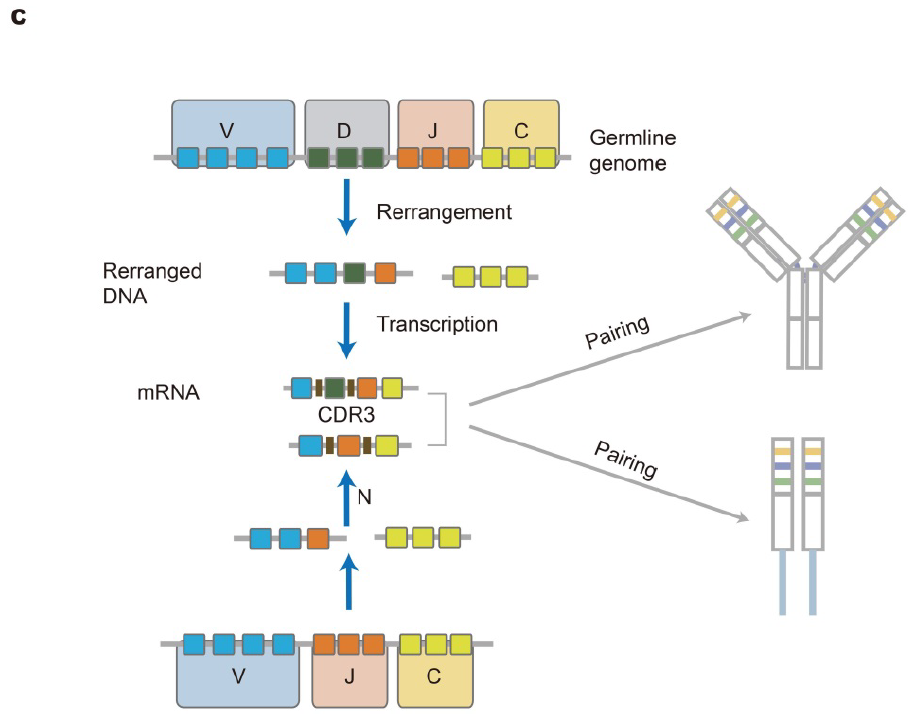
图2 淋巴细胞受体库多样性的机制[1]
注:在淋巴细胞发育过程中,BCR重链或TCRβ(δ)链进行VDJ基因的重排,而IgL链或α(γ)链在重排中缺少D基因;然后,将重新排列的V-DJ或V-J序列连接到C基因片段。最后,两条独立的链被组装成独特的受体蛋白。胚系基因V(D)JC经历核苷酸的重排、插入和删除,导致受体库的多样性
PART 02
TCR重排和基因结构
TCR 是由 α 和 β 链 (TCRαβ) 或 γ 链和 δ 链 (TCRγδ) 组合形成的跨膜糖蛋白异二聚体。它们仅在 T 细胞中表达,其中大多数 (95%) 为 TCRαβ 异二聚体。TCRα 和 TCRβ 胞外结构域由肽段识别至关重要的可变结构域(Vα 和 Vβ)和恒定结构域(Cα 和 Cβ)组成。
编码α链和β链(分别为 TRA 和 TRB)的基因由多个不连续的片段组成:TRA 和 TRB 的可变区 (V) 和连接区 (J),以及 TRB 的多样性 (D) 片段。TCR 基因经历 V(D)J 重组:α 链的 V 和 J 片段以及 β 链的 V、D 和 J 片段间重组,形成功能可变的编码序列。然后,该序列与 C 片段重组,并最终转录为功能性 TRA 或 TRB 链转录本。TCRα和TCRβ链的结构中都包含三个高变区,分别称为互补决定区1、2和3(CDR1、CDR2、CDR3)。CDR1 和 CDR2 区域由 V 段胚系基因编码,CDR3区由连接区V(D)J基因编码,片段间伴有核苷酸(N)随机插入/删除,为该区域提供高度的变异性。这些过程最终产生由特定α和β链组成的 TCR,这些链决定了受体的特异性(图3)。
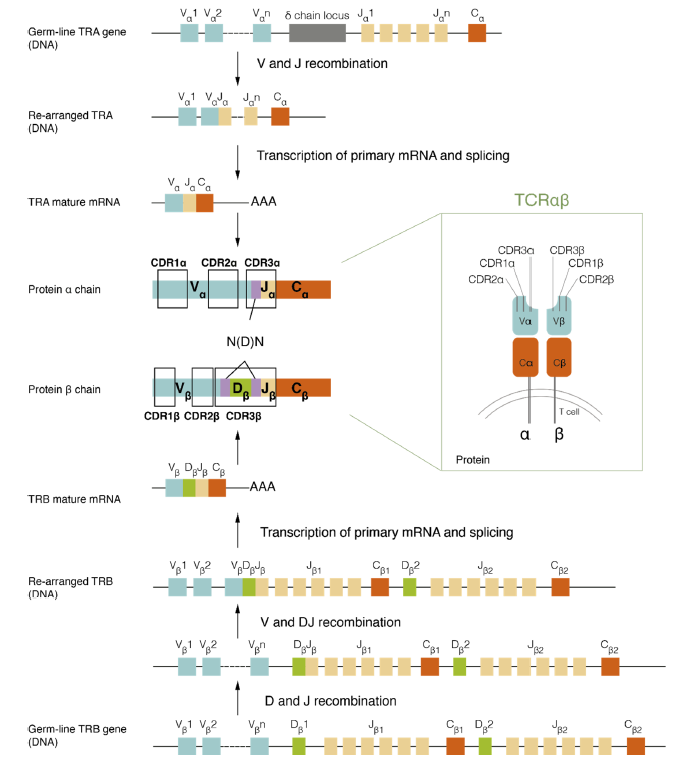
图3 TCR的结构组成[2]
PART 03
BCR重排和基因结构
人类的抗体是蛋白质复合物,其基本单位是一对二硫键连接的重链蛋白和一条对应的轻链(图3)。重链和轻链的N末端区域的序列高度可变,并且是抗体的抗原结合部位。蛋白质的C末端区域被称为恒定区域。轻链有两种类型,kappa (IgK) 和lambda (IgL),而重链(IgH)有五种主要的类型(IgM、IgD、IgG、IgA和IgE),其中IgG有四种亚型,IgA有两种亚型。
为了识别不同的抗原,在每个B细胞中,Ig基因经历两次多样化过程,包括抗原非依赖性和抗原依赖性过程,最后形成总的BCR抗体库。

非抗原依赖的胚系重排过程
在暴露于抗原之前,V(D)J重组发生在网膜、胎儿肝脏或成人骨髓中的B细胞发育过程中,IgV外显子重排组装形成一级抗体库。在这种不依赖抗原的过程中,胚系可变区(V)、高变区(D)和连接区(J)基因片段以有序的方式在Ig基因座中重排组装而成,首先将D和J片段连接在一起,然后选择V片段连接到新产生的D-J产物,从可变(V)、高变区(D)和连接区(J)基因片段胚系序列中组装编码其免疫球蛋白重链的抗原结合区的基因。在连接过程中,对基因片段的末端进行核酸外切酶消化,并在片段连接处添加非模板随机碱基(N)。初级BCR库的多样性来自V/D/J基因片段的数量,以及连接点的插入和删除(indel)突变。在IgV结构域中,该序列可进一步分为互补决定区(CDR)和框架区(FR)。FR和CDR1/2来自V基因片段,而CDR3覆盖包含V-D-J连接的序列(图3)。因此,CDR3是最多样化的序列,通常在抗原识别过程中发挥最重要的作用,但由可变基因片段编码的另外两个结构域(CDR1和CDR2)也可以显著促进抗原特异性。从肽序列方面来看,CDR3有以下几个特征,即,以半胱氨酸(C)开始,以苯丙氨酸(F)或色氨酸(W)结束。 在表达功能性IgH后,κ或λ免疫球蛋白轻链基因座上发生类似的V和J片段重排。
在暴露于抗原之前,个体B细胞抗体库的遗传复杂性来源于V、(D)和J片段的重排组合,编码抗体重链或轻链的CDR3区域片段连接处的多样化,以及重链和轻链的配对组合。早期对这些机制可能产生的不同抗体的数量的粗略估计为大于1011。这个数字应该是被低估了,因为抗原刺激下B细胞激活体细胞超突变为抗体序列多样化提供了更多可能,该过程是在整个重排的V(D)J序列中产生新的点突变,形成克隆后代,其抗体可能对抗原具有更强的结合亲和力;另外受体编辑等罕见过程通过V基因片段的二次重排和替换,为抗体库的生成提供了进一步的可能。在人类群体水平上,抗体库的更多遗传多样性源于免疫球蛋白等位基因突变和免疫球蛋白基因座内扩增或删除的拷贝数变异。
V基因片段的遗传结构包括编码大部分前导肽的上游外显子,其次是短内含子、编码其余前导肽和V片段的第二外显子。在未重排的人类胚系基因座中,V、D和J区域彼此相隔数千碱基,由单个外显子(κ或λ)或几个外显子编码的恒定区域(重链不同类型)位于J片段下游数千碱基处,除λ基因座外,其中一些J片段散布在最上游的恒定区域之间。在重链和轻链基因座重排过程中产生长度约为400个碱基紧凑的V(D)J基因。
根据IMGT数据库(www.IMGT.org)报道,14q32人类IGH基因座的序列分析表明,人类具有123-129个IGHV基因片段,其中43-46个能够形成功能重排;27个IGHD基因片段(23个功能片段);以及9个IGHJ片段(6个功能性片段)。2p11的IGK基因座在大多数个体中编码76个IGKV片段(31-36个功能性片段)和5个IGKJ基因片段(5个功能性片段),而22q11的IGL基因座包含73-74个IGLV片段(29-33个功能性片段)和7-11个IGLJ片段(4-5个功能性片段)。随着技术的发展,识别Ig基因新变异的步伐已经加快,这表明目前分析的变异只代表了所有人类群体中可能存在的总变异的一部分。
抗原依赖的二次多样化
抗原刺激后,BCR经历另一次多样化,包括IgH类开关重组(CSR)和IgV体细胞超突变(SHM),这两种突变都是由激活诱导的胞苷脱氨酶(AID)引发。在CSR中,BCR可以从IgM转换到另一种同种型,包括IgG、IgA或IgE。而在SHM中,AID可以在IgV外显子处启动突变或INDEL。表达突变多样性BCR的B细胞在生发中心的次级淋巴结构中进行亲和力的选择,该过程为亲和力成熟的过程。伴随着这些抗原依赖性过程,幼稚B细胞进一步分化为分泌抗体的浆细胞或记忆B细胞。此时,B细胞库表达高级抗体库,其中包含不同类别的高效抗体。人类抗体的体细胞超突变主要发生在次级淋巴组织的特殊微环境中,其中B细胞可以接触抗原、特异树突状细胞、基质细胞和T细胞,以及通过可溶性介质和细胞-细胞接触刺激B细胞。在急性抗原刺激(如流感疫苗接种)后观察到的人浆母细胞在IGHV片段中通常显示出5%-15%的突变水平,而记忆B细胞显示出略低的突变水平。在特殊的情况下,如HIV感染者长期暴露于病毒抗原免疫系统紊乱时,可以观察到更高的突变水平(超过30%)。突变事件主要集于前导序列的上游至V(D)J重排的区域,并在将J片段与恒定区域之间的内含子区中逐渐变少。
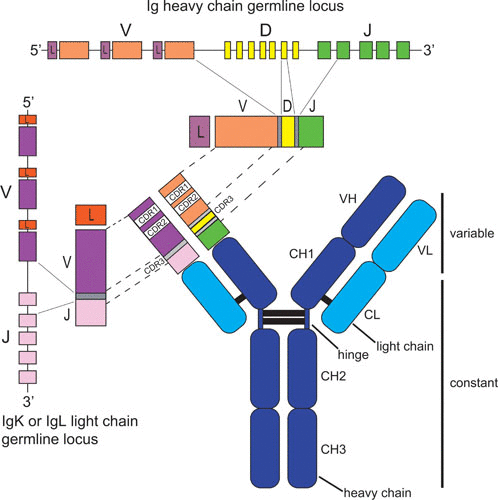
图4 抗体结构和基因编码[3]
注:免疫球蛋白重链基因座的胚系(未重排)基因组DNA结构如图顶部所示,显示了 V、D 和 J 基因片段的串联排列(非等比例缩放)。胚系κ或λ轻链基因座位于左侧,具有未重排的V和J片段。胚系DNA的逐步重排使重链D和J基因片段的连接,随后V片段与D-J产物的连接,产生编码重链可变区的DNA。在重排过程中,基因片段的末端经过核酸外切酶消化,并在片段末端添加随机的非模板碱基,以在编码互补决定区3(CDR3)的VDJ连接区产生额外的序列多样性,其通常是抗体重链中对抗原特异性影响最大的区域。在轻链基因座中发生了类似的V和J基因重排过程,VJ连接多样化,从而产生重排的轻链基因。重链和轻链的恒定区(重链结构域CH1、CH2和CH3,轻链结构域CL)由下游外显子编码,这些外显子通过mRNA剪接与重排的V(D)J基因连接。抗体结构中蛋白链连接的二硫键用黑线显示。
本篇内容给大家介绍了什么是免疫组库测序,详细介绍了BCR/TCR基因重排的过程,下篇将为大家介绍免疫组库测序的方法,敬请期待。
参考文献:
[1] Liu H, Pan W, Tang C, Tang Y, Wu H, Yoshimura A, Deng Y, He N, Li S. The methods and advances of adaptive immune receptors repertoire sequencing. Theranostics. 2021 Aug 19;11(18):8945-8963. doi: 10.7150/thno.61390. PMID: 34522220; PMCID: PMC8419057.
[2] Aran A, Garrigós L, Curigliano G, Cortés J, Martí M. Evaluation of the TCR Repertoire as a Predictive and Prognostic Biomarker in Cancer: Diversity or Clonality? Cancers (Basel). 2022 Mar 31;14(7):1771. doi: 10.3390/cancers14071771. PMID: 35406543; PMCID: PMC8996954.
[3] Boyd SD, Joshi SA. High-Throughput DNA Sequencing Analysis of Antibody Repertoires. Microbiol Spectr. 2014 Oct;2(5). doi: 10.1128/microbiolspec.AID-0017-2014. PMID: 26104353.
[4] Li N, Yuan J, Tian W, Meng L, Liu Y. T-cell receptor repertoire analysis for the diagnosis and treatment of solid tumor: A methodology and clinical applications. Cancer Commun (Lond). 2020 Oct;40(10):473-483. doi: 10.1002/cac2.12074. Epub 2020 Jul 17. PMID: 32677768; PMCID: PMC7571402.
[5] Joshi K, Milighetti M, Chain BM. Application of T cell receptor (TCR) repertoire analysis for the advancement of cancer immunotherapy. Curr Opin Immunol. 2022 Feb;74:1-8. doi: 10.1016/j.coi.2021.07.006. Epub 2021 Aug 25. PMID: 34454284.
[6] 12 Zhang S, Yang T, Liu X, Yang J, Zheng X. Antibody repertoire sequencing analysis. Acta Biochim Biophys Sin (Shanghai). 2022 May 25;54(6):864-873. doi: 10.3724/abbs.2022062. PMID: 35713313; PMCID: PMC9828323.
[7] Nixon AB, Schalper KA, Jacobs I, Potluri S, Wang IM, Fleener C. Peripheral immune-based biomarkers in cancer immunotherapy: can we realize their predictive potential? J Immunother Cancer. 2019 Nov 27;7(1):325. doi: 10.1186/s40425-019-0799-2. PMID: 31775882; PMCID: PMC6880594.
[8] Schrama D, Ritter C, Becker JC. T cell receptor repertoire usage in cancer as a surrogate marker for immune responses. Semin Immunopathol. 2017 Apr;39(3):255-268. doi: 10.1007/s00281-016-0614-9. Epub 2017 Jan 10. PMID: 28074285.
[9] Porciello N, Franzese O, D'Ambrosio L, Palermo B, Nisticò P. T-cell repertoire diversity: friend or foe for protective antitumor response? J Exp Clin Cancer Res. 2022 Dec 22;41(1):356. doi: 10.1186/s13046-022-02566-0. PMID: 36550555; PMCID: PMC9773533.